El esclerénquima es uno de los dos tejidos de sostén que poseen los vegetales. Su nombre deriva de dos palabras griegas:
Comprende complejos de células que otorgan a la planta resistencia a los estiramientos,
torceduras, pesos y presiones. En algunos casos la fortaleza y elasticidad de las paredes
secundarias de ciertas células sirven como medio defensivo, ya que otorgan a la planta resistencia contra las partes bucales, garfios y ovipositores de los insectos.
Las células esclerenquimáticas se diferencian de las colenquimáticas en que poseen paredes
secundarias generalmente lignificadas y en que, cuando adultas, carecen frecuentemente de
protoplasma.
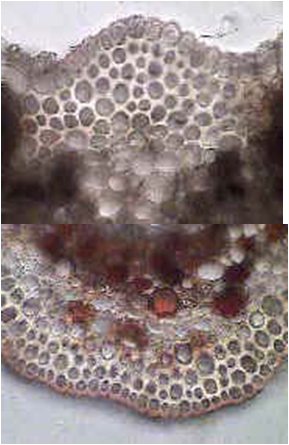
Corte transversal de tallo

Las células del esclerénquima deben sus propiedades a la
celulosa, hemicelulosas, y además hasta 30% de lignina. La lignificación se produce desde afuera hacia adentro, comienza en la laminilla media y pared primaria; la capa S3 a veces es solamente celulósica.
Se han propuesto una variedad de sistemas para la clasificación de las células
esclerenquimáticas. Aquí consideramos dos tipos básicos, que se diferencian por la forma de las células:
Las esclereidas y fibras pueden quedar asociadas a tejidos primarios o secundarios, de manera que ontogenéticamente se desarrollan a partir de diferentes
meristemas primarios derivados: procámbium, meristema fundamental e incluso la protodermis, o de los meristemas secundarios: cámbium y felógenoesclereidas y fibras. Cuando es difícil ubicar una célula en una u otra categoría, se puede usar el término fibroesclereidapared secundaria, que presentaCuando se originan durante el crecimiento primario, las células esclerenquimáticas crecen primero por crecimiento simplástico, coordinado con el de las células vecinas en división, y sin modificar los contactos celulares y las comunicaciones intercelulares. Durante este período las fibras pueden volverse multinucleadas como resultado de mitosis repetidas no acompañadas de citocinesis.
Luego las fibras, y las esclereidas ramificadas aumentan en longitud por crecimiento apical
intrusivo, los extremos se abren paso entre las células vecinas o entre los espacios intercelulares. Si los extremos son obstruidos por otras células se curvan o se bifurcan
La porción media que ya ha completado su alargamiento desarrolla paredes secundarias, pero las paredes de los extremos permanecen delgadas
La pared se forma de una sucesión de tubos que se van alargando. Las paredes secundarias son generalmente flojas y relajadas durante su formación, y se presentan a veces separadas de la pared primaria
Esquemas mostrando el crecimiento en longitud de una fibra y al mismo
tiempo el depósito de las sucesivas capas de la pared secundaria

Esclereidas
Se definen como esclereidas a las células del esclerénquima de forma muy variada,
frecuentemente cortas.
Pueden encontrarse en diferentes órganos de la planta, incorporadas a tejidos diversos, primarioso secundarios. Se las halla solitarias o agrupadas, pero nunca formando cordones como las fibras.
Los carozos de las drupas y las cubiertas de muchas semillas deben su dureza a que están
constituidos por esclereidas.
Braquiesclereidas en tallo de
Hoya carnosa, flor de nácar (Dicot

scleros (duro) y enchyma (sustancia o infusión). Es un tejido elástico, es decir que puede ser deformado por tensión o presión, pero retoma su forma original cuando la fuerza desaparece.